
Eicosanoides. su funcion en la inflamación
Trabajo enviado por:
María Azucena Mendoza Fernández
Indice
1. Introducción
2. Origen de los Eicosanoides
3. El ácido araquidónico: precursor de las prostaglandinas, los tromboxanos y los leucotrienos.
4. Biosíntesis de las Prostaglandinas y de los tromboxanos
5. Biosíntesis de Leucotrienos
6. Células que producen eicosanoides
7. Eicosanoides. Mediadores químicos de la inflamación
8. Bibliografía
1. Introducción
Los eicosanoides son sustancias fisiológicamente activas cuya función y efectos en los sistemas biológicos son muy amplios y aún no han sido aclarados en su totalidad. Actúan como potentes reguladores intracelulares participando en gran medida en los procesos inflamatorios y en la respuesta inmune.
El primer eicosanoide estudiado fue la prostaglandina, en 1933 Goldblatt en Inglaterra y Euler en Suecia descubrieron propiedades en el líquido seminal y en el plasma sanguíneo. En 1957 Bergstrón y Sjöval aislaron prostaglandinas de la próstata de donde surgió el nombre de Prostaglandina, y determinaron que estas sustancias son ácidos grasos poliinsaturados, también se estudió su síntesis enzimática la cual consta por lo menos de dos sistemas subsecuentes. Sus efectos no se conocen todavía con precisión pero ya se establecieron dos características generales: 1) muchas células blanco controladas por prostaglandinas contienen proteínas receptoras específicas de membrana, mismas que se fijan a prostaglandinas individuales y 2) las prostaglandinas incrementan o reducen, dependiendo del tejido, las concentraciones celulares de AMP cíclico, GMP cíclico o ambos.
El tromboxano se identificó en 1975, se caracterizó por ser un potente agente iniciador de la agregación plaquetaria. Se identifico que éste se sintetizaba en las plaquetas y en otros tejidos.
Los leucotrienos recibieron este nombre por producirse en los leucocitos y además porque contienen tres dobles enlaces conjugados, éstos son constrictores del músculo liso, y se relacionan con las dificultades respiratorias de los asmáticos.
El estudio de la síntesis de los eicosanoides y su relación con los procesos Fisiológicos normales y anormales en el organismo ha sido de gran valor, sobre todo para la creación de nuevos productos terapéuticos que controlan la inflamación y el dolor.
Los eicosanoides son derivados de ácidos grasos poliinsaturados de veinte átomos de carbono 1,2 (de ahí les viene su nombre ya que Eikosi- es un prefijo griego que significa veinte)2,6 y que poseen 3, 4 ó 5 dobles enlaces. También se les conoce como prostanoides debido a que se consideran derivados hipotéticos hidrocarbonados de prostana y trombana y a sus correspondientes ácidos prostanóico y trombanóico 4,6.
Los precursores poliinsaturados de los eicosanoides son los ácidos eicosaenoicos:
- El ácido gamma-linoleico ( ácido-homo-gamma-linoleico)
- El ácido araquidónico
- El ácido eicosapentaenoico 1,3,5.
Los ácidos eicosaenoicos se sintetizan a partir de precursores conocidos como ácidos grasos esenciales, llamados así porque no son sintetizados en el organismo, por lo que deben ser ingeridos a través de la dieta. Estos ácidos grasos esenciales se absorben a través del intestino y por medio de la sangre son llevados hasta el hígado9, donde se sintetizarán los ácidos eicosaenoicos, los cuales se almacenan en la bicapa lipídica de las membranas celulares. Si la dieta es rica en vegetales el organismo se enriquecerá de ácido dihomo-gamma-linoleico, si es rica en carnes rojas se enriquecerá de ácido araquidónico y si es rica en pescado de ácido icosapentaenoico3. Por ejemplo el ácido araquidónico es sintetizado a partir de ácido linoleico, siendo este un ácido graso esencial que se aporta a la dieta a partir de las carnes en cantidades de al menos 10 g por día9.
Para que de inicio la síntesis de los eicosanoides se necesita de ciertos estímulos químicos (hormonas o neurotransmisores), físicos (calor, corriente eléctrica)3,5, de hipoxia, etc. Una vez que los ácidos eicosaenoicos reciben alguno de estos estímulos, son liberados de los fosfolípidos de la membrana celular por la acción de diversas enzimas, como son: las fosfolipasas, ciclooxigenasas, lipooxigenasas y por el citocromo P-4501,5. Así se forman los eicosanoides los cuales están constituidos por: Prostaciclinas, Prostaglandinas, Tromboxanos, Leucotrienos, lipoximas y otros ácidos grasos hidroxilados8.
3. El ácido araquidónico: precursor de las prostaglandinas, los tromboxanos y los leucotrienos.
El ácido araquidónico (o ácido icosatetraenoico llamado así porque posee cuatro dobles ligaduras) es el ácido eicosaenoíco más abundante en el hombre1. Es además el precursor más importante de los eicosanoides. Se sintetiza a partir del ácido linoleico por elongación y desaminación en el hígado y se acumula en las membranas de las células eterificando al C(2) del glicerol del fosfatidilinositol y de otros fosfolípidos6.
La producción de ácido araquidónico está controlada por la velocidad de la liberación de estos fosfolípidos a través de tres rutas alternativas:
- la fosfolipasa A2 hidroliza los grupos acilo en C(2) de los fosfolípidos,
- la fosfolipasa C hidroliza específicamente el grupo de cabeza de fosfatidilinositol a fin de rendir un 1,2-diacilglicerol, que se fosforila por la diacil-glicerol quinasa a ácido fosfatídico que es un sustrato de la fosfolipasa A,
- el 1,2-diacilglicerol puede hidrolizarse también directamente por la diacilglicerol lipasa 5,6. (figura 1)
A partir de esta etapa las distintas vías de la llamada "cascada del ácido araquidónico" originan un amplio espectro de compuestos biológicamente activos. La preponderancia de las distintas vías depende del tejido en cuestión, ya que cada sistema celular le ofrece al ácido araquidónico distintos tipos de enzimas para su transformación3. Se conocen cinco vías de oxidación del ácido araquidónico, en esta monografía solo se describen las vías que conducen a formar sustancias de importancia en los procesos inflamativos, estas son la ruta cíclica del metabolismo del araquidonato que origina las prostaglandinas y los tromboxanos y la ruta lineal que origina los leucotrienos.
Figura 1. Tres Rutas de liberación del Ácido araquidónico a partir de fosfolípidos6
4. Biosíntesis de las Prostaglandinas y de los tromboxanos
La ruta cíclica del ácido araquidónico (figura 2) se inicia con la participación de la enzima sintetasa del endoperóxido la cual tiene dos componentes catalíticos :
- La enzima ciclooxigenasa que convierte al ácido araquidónico en endopoeróxido cíclico (PGG2) por introducción de oxígeno molecular y formación de ciclopentano3,10. La PGG2 tiene un puente endoperóxido (-O-O-) entre el anillo del ciclopentano y un grupo de hidroxiperóxido (-OOH).
- A partir de PGG2 se forma el segundo endoperóxido cíclico (PGH2) por acción de la enzima peroxidasa, la cual reduce el grupo hidroperóxido del C15 a grupo hidroxilo (-OH). La PGH2 conserva su función de endoperóxido (-O-O-)3,6,7,10.
Como producto de esta reacción se originan agentes oxidantes que regulan la acción de la ciclooxigenasa y además poseen importancia en el proceso inflamatorio3,6,7,10,11.
La PGH2 es el precursor inmediato de todas las prostaglandinas de la serie 2, las prostaciclinas y los tromboxanos6.
Las PGG2 y PGH2 poseen vida media muy corta y son metabolizadas rápidamente para dar origen a sus productos, cuatro de ellos son prostaglandinas: PGE2 (soluble en éter), PGF2 (soluble en tampón de fosfato), PGD2 y PGI2, 6-oxo-PGF1 alfa, tromboxano A2, (un compuesto inestable) y Tromboxano B23,6. La etapa de transformación del ácido araquidónico a PGH2 es común en todos los sistemas celulares El metabolismo posterior del PGH2 depende de: la característica de cada célula particular, de las enzimas que contenga y del balance en cofactores 1,3,6.
Las prostaglandinas son clasificadas en nueve grupos que se distinguen por los sustituyentes en un anillo de 5 átomos de C. Las prostaglandinas de la serie F se clasifican de acuerdo a la configuración del grupo –OH en el C9 como alfa y beta. Las G, H e I no difieren en cuanto a los sustituyentes del anillo, pero si en el grupo R2, la prostaglandina G tiene el grupo 15-S-hidroxiperóxido en lugar del 15-S-hidroxi, que está en la prostaglandina H. El número después del nombre de la prostaglandina enseña la cantidad de enlaces dobles en las cadenas R1 y R2 11.
Los tromboxanos de configuración natural se dividen en dos grupos: Tromboxanos A (TxA) y tromboxanos B (TxB)11.
5. Biosíntesis de Leucotrienos
La ruta lineal del ácido araquidónico (figura 3) se inicia en la reacción de conversión del araquidonato catalizada por la enzima lipooxigenasa en las posiciones 5,12,15 para formar ácidos hidroperoxieicosatetraenoicos (HPETEs), sustancias que por si mismas no son mediadores fisiológicos. La reacción consiste en la separación de hidrógeno de una unidad de 1,4-pentadieno, la adición de oxígeno y la adición de nuevo del átomo de hidrógeno3,6.
La enzima lipooxigenasa convierte al ácido araquidónico en un hidroperóxido llamado ácido eicosatetraenoíco (5-HPETE). El intermediario 5-HPETE pierde agua para formar en primer lugar un epóxido insaturado el Leucotrieno A4 (LTA4), el subíndice indica el número de enlaces dobles C-C en la molécula así como la serie a la que pertenece el Leucotrieno. A continuación la enzima glutatión-S-transferasa catatiza la adición del grupo sulfhidrilo del glutatión al epóxido formando el primero de los peptidoleucotrienos, el Leucotrieno C4 (LTC4). La enzima gamma-glutamiltransferasa separa ácido glutámico convirtiendo LTC4 en Leucotrieno D4 ( LTD4), éste último se convierte en Leucotrieno E4 (LTE4) por la acción de la enzima dipeptidasa que elimina glicina. LTA4 puede convertirse también en Leucotrieno B4 (LTB4) por la acción de la enzima Leucotrieno A4 hidrolasa3,6,12 . El intermediario del LTA4 el 5-HPETE también puede convertirse en ácido 5-hidroxieicosatetraenoíco (5-HETE)12. Se conocen otras lipooxigenasas como la 11, la 12 y la l5 lipooxigenasas las cuales originan 11-HPETE, 12-HPETE y 15-HPETE respectivamente, estas a su vez pueden formar 11-HETE, 12-HETE Y 15-HETE . Algunos de estos derivados HPETE y HETE pueden ser antagonistas o protagonistas de la actividad de los Leucotrienos, Prostaglandinas o ambos. Otros pueden ser precursores de sustancias fisiológicamente activas ó funcionar como sustancias quimiotácticas3,6,7,1

Figura2. Ruta cíclica del ácido araquidónico. La ramificación de la ruta conduce a la
Formación de prostaglandinas, prostacilinas y tromboxano.6
6. Células que producen eicosanoides
Las plaquetas son ricas en la enzima tromboxano sintetasa y producen una cantidad elevada de tromboxano A2.
Las células del endotelio vascular son ricas en la enzima prostaciclina sintetasa formando PGI2. Las PGE2, PGF2 Y PGD2 se encuentran difundidas en casi todas las células del cuerpo la excepción son los glóbulos rojos y los linfocitos. Las células del músculo cardiaco producen PGI2, PGE2 y PGF2 alfa en cantidades más ó menos iguales6. Las células del encéfalo sintetizan PGD2, las células endoteliales del hígado sintetizan PGI2. Las células endocrinas sintetizan PGE1 y PGE2. PGF2 alfa se produce en el útero, aquí influye sobre el cuerpo amarillo. La PGI2 tiene dos tipos de receptores en plaquetas4,14 .
Los leucocitos forman Leucotrienos (LT). Por ejemplo los polimorfonuclares como los neutrófilos y los eosinófilos producen LTB4, y los basófilos LTC4, LTD4 y LTE4. Los mastocitos producen los mismos LT que los basófilos. Las células del pulmón también los producen, y tienen una participación importante como agentes broncoconstrictores3,14.
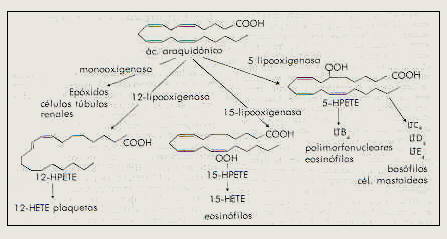
Figura 3. Inicio de la ruta cíclica y la ruta lineal del metabolismo del ácido araquidónico6
7. Eicosanoides. Mediadores químicos de la inflamación
La reacción inflamatoria es un mecanismo de defensa natural que provee el medio por el cual los factores protectores como los anticuerpos, el complemento y las células fagocíticas (polimorfonucleares y mononucleares), que se encuentran localizados normalmente en la sangre, puedan penetrar al tejido y ganar acceso a los sitios de invasión de elementos extraños. La inflamación es considerada como un medio destinado a focalizar los mecanismos inmunológicos protectores en una región localizada dentro del tejido.
En la inflamación aguda los vasos sanguíneos se dilatan, por esta razón se presenta rubor y calor en el área, el edema se produce por escape de fluidos y de células al tejido extravascular. El dolor y la perdida de función se deben a la presión ejercida en las terminales nerviosas por la acumulación de líquido extravascular y a la liberación de mediadores químicos.
Un mediador químico de la inflamación debe cumplir con los siguientes criterios:
- el mediador debe ser detectable en el sitio de la inflamación en el momento oportuno y en cantidades adecuadas para realizar el efecto en consideración.
- el mediador, cuando se administre en concentraciones del orden de las encontradas en la lesión, debe producir los efectos observados y no otros.
- el efecto se debe atenuar o prevenir con agentes bloqueadores específicos o antagonistas del mediador propuesto.
- la prevención de la liberación del mediador debiera abolir o prever el efecto, y agentes o procedimientos que prevean la degradación o desaparición del mediador debieran prolongar o potenciar el efecto.
En base a los criterios expuestos se consideran mediadores químicos de la inflamación: la histamina, la bradikinina, el factor activador de plaquetas y los eicosanoides prostaglandinas (PG), tromboxanos (Tx) y leucotrienos (LT) 1,2,3,7,8,11,13,14.
Las PG, los Tx y los LT son considerados como hormonas locales5,4,3,7 debido a que no existen depósitos de ellos en las células, sino que se sintetizan y liberan localmente según la demanda, se metabolizan con rapidez por lo que son detectables solo por periodos cortos3. Pueden actuar como mediadores en casi todos los procesos de la inflamación aguda1.3.13 y se reconoce que cualquier proceso que conduce a la ruptura de la membrana lleva a la liberación de PG3.
La vasodilatación y el enrojecimiento son causados por la liberación de PGI2, PGE2, PGE1 y PGD2, y su efecto es antagonizado por TxA2. Las PG vasodilatadoras poseen la habilidad de aumentar el edema producido por bradikinina e histamina 1,3, dilatan arteriolas dando como resultado una dilatación pasiva en las vénulas por un incremento en la presión hidrostática en la luz de la vénula. Así junto con la bradikinina e histamina aumentan la cantidad de proteína plasmática exudada. Las PG mas importantes en este fenómeno son la PGE2 y PGI2. La PGE1, E2 e I2 producen dolor, siendo la mas efectiva PGI2 debido a su capacidad de elevar los niveles intracelulares de AMP cíclico3,11,13. PGE2 interacciona con la IL-1 Iinterleucina 1) y FNT (factor de necrosis tumoral) para producir la fiebre que se observa en las respuestas inflamatorias sistémicas que aparecen en las infecciones 13. Al inicio de la inflamación aguda las células del tejido dañado son las productoras de prostanoides, esto en base a que los fagocitos PMN ( neutrófilos y eosinófilos) no son productores eficientes de prostaglandinas3,12,14.
El LTB4 es un mediador natural de la inflamación. Los exudados que se producen en el inicio del proceso inflamatorio son ricos en leucocitos formadores de LTB4, posteriormente interactúa con las PG, las cuales ejercen su acción causando aumento en la permeabilidad vascular y formación local de edema. De esta forma las PG y el LTB4 pueden actuar en forma secuencial y en combinación produciendo la permeabilidad vascular y la infiltración celular que caracteriza la reapuesta inflamatoria. Los LT participan en forma indirecta en la inflamación ejerciendo diversos efectos sobre los leucocitos:
- expresión de receptores de superficie.
- aumento de la expresión de los receptores de C3b en eosinófilos.
- incrementando los niveles de AMP-cíclico intracelular.
- estimulación en la captación de Ca++ extracelular.
- estimulación en la captación de D-glucosa.
Los leucotrienos C4, D4 y E4 son los principales constituyentes de la sustancia de liberación lenta de la anafilaxis (SRS-A), funcionan como importantes mediadores químicos de la inflamación por su acción en el lecho microvascular. Estos producen exudación del plasma de las vénulas postcapilares con una eficacia mayor que la histamina. El aumento de la permeabilidad producido por estos Leucotrieneos es directo sobre las células endoteliales, ocurre rápidamente y no requiere la presencia de histamina, prostaglandinas ni leucocitos
PMN. La presencia única de Leucotrienos SRS-A produce edema, contribuyendo a uno de los signos primordiales del proceso inflamatorio.3,11,13 (figura 4) En la tabla 1 se puede observar la contribución de las PG, los Tx y los LT al proceso inflamatorio. Los esteroides con actividad anti-inflamatoria inhiben la liberación del ácido araquidónico de los fosfolípidos de membrana, al inhibir la acción de la fosfolipasa A2 por liberación de un péptido llamado lipomodulina, de esta manera impide la formación de los eicosanoides. La aspirina inhibe la transformación del araquidonato en PG y Tx.
La participación de las PG, Tx y LT en la inflamación es relevante, no solo para mediar la inflamación sino también para ejercer un balance por sus efectos antagonistas. El estudio de los eicosanoides y de su relación con los procesos biológicos es hoy en día motivo de estudio, ya que a medida que se van comprendiendo mejor los mecanismos de acción de las PG, Tx y LT se mejoran los agentes terapéuticos anti-inflamatorios.
Figura 4. Proceso Inflamatorio mediado por Leucotrienos3,13
| TABLA I EFECTO DE LOS EICOSANOIDES EN LA INFLAMACIÓN PROSTAGLANDINAS PGE2 PGI2 TROMBOXANO TxA2 LEUCOTRIENOS LTB4 SRS-A Vasodilatación vasodilatación vasoconstrictor quimiotaxis vasoconstrictor Edema edema agregatorio degranulación edema Dolor dolor broncoconstrictor extravasación Broncoconstrictor antiagregatorio antiagregatorio adhesión Aumento de la Permeabilidad Vascular Fiebre edema transvasación Infiltración Celular Migración diapédesis
|
8. Bibliografía
- www.drscope.com/pac/anestesia-1/b4/an1b4-p21.htm. Eicosanoides
- campus.usa/.es/-dbbm/modmol/modmol/03/mm03t01.htm. Lípidos
- Margni, Ricardo A.; Inmunología e Inmunoquímica; cuarta edición; Buenos Aires Argentina. 1989;pp 325-339.
- Ilyiná A.D., "Las prostaglandinas y tromboxanos. Su función como biorreguladores celulares. Parte I" Bioquimia 1995; 78:243-247
- www.lafacu.com/apuntes/medicina/medi_celuz/ Mediadores Celulares: Eicosanoides
- Voet Donald y Judith; Bioquímica; editorial Omega; Barcelona 1992;pp 704-711
- Matheus, Christopher K, Van Holde K.E.; Bioquímica; McGraw Hill.Interamericana segunda Edición; España 1988; pp 764-769
- Díaz Argüelles Virginia; "Suplementación enteral de los ácidos grasos esenciales en RN pretérmino"; Rev. Cubana Pediatr 2001; 73(1) :34-42
- www.iqb.es/cbasicas/Bioquim/Cap7/C7a10_01.htm Curso de Bioquímica
- Lehninger, Albert L; Bioquimica; Ed. Omega, Barcelona; pp 306-307,682,689
- Stites P, Daniel, Stobo D, John; Inmunología Básica y Clínica; octava edición, El Manual Moderno. México 1966 pp214-215
- Bohinski, Robert C; Bioquímica; Quinta Edición; Wilmington Delaware EUA 1991; pp 45, 418, 422, 432-35, 631,649
- bvs.sld.cu/revistas/est/vol35_2_98/est04298.htm Biosíntesis de Eicosaenoides
- Abbas, Kabul A,Inmunología Celular y Molecular,Tercera edición, Editorial McGraw Hill, Aravaca Madrid 1999, pp 431-432
María Azucena Mendoza Fernández. Facultad de Química Farmacobiológica. Universidad Veracruzana. Universidad Autónoma de Tamaulipas. Maestría en Análisis Clínicos. Bioquímica General. México.
Resumen
Los eicosanoides son sintetizados en la membrana celular de casi todas las células del organismo, a excepción de los glóbulos rojos, como respuesta a un estímulo que puede ser químico o físico. Su síntesis está regulada por enzimas específicas para cada célula en particular, de tal forma que no todas las células producen el mismo tipo de eicosanoides. Su participación en los procesos biológicos es muy amplia, pero merece especial atención el proceso inflamatorio, ya que éstos median muchos de los fenómenos que ocurren en este proceso. Los eicosanoides que están relacionados con la inflamación son las prostaglandinas, los leucotrienos y los tromboxanos. Estos son considerados como "hormonas locales" debido a que al igual que las hormonas, ejercen efectos fisiológicos importantes actuando en concentraciones extremadamente bajas y en un área limitada por el mismo proceso. No se encuentran concentraciones de ellos en los tejidos a menos que éste sea dañado, por lo que su producción es una respuesta a la irritación del tejido. Se sintetizan en su mayor parte a partir del ácido araquidónico, el cual es liberado de la membrana celular por acción enzimática originando la "cascada del araquidonato". La ruta cíclica de la cascada da origen a las prostaglandinas y los tromboxanos y la ruta lineal a los leucotrienos, Sus funciones en la inflamación es como agonistas y antagonistas de tal manera que modulan la respuesta inflamatoria.
Palabras clave: eicosanoides, prostaglandinas, leucotrienos, tromboxanos, inflamación.
Trabajo enviado por:
María Azucena Mendoza Fernández
azulzuly[arroba]hotmail.com